Central nervous system (CNS) is formed in week 3 of development, during which time the neural plate develops. The neural plate consists of neuroectoderm and becomes the neural tube, which gives rise to the brain and spinal cord.
Peripheral nervous system (PNS) is derived from three sources:
Neural crest cells
Neural tube, which gives rise to all preganglionic autonomic nerves (sympathetic and parasympathetic) and all nerves (alpha-motoneurons and gamma-motoneurons) that innervate skeletal muscles
Mesoderm, which gives rise to the dura mater and to connective tissue investments of peripheral nerve fibers (endoneurium, perineurium, and epineurium)
Development of the Neural Tube
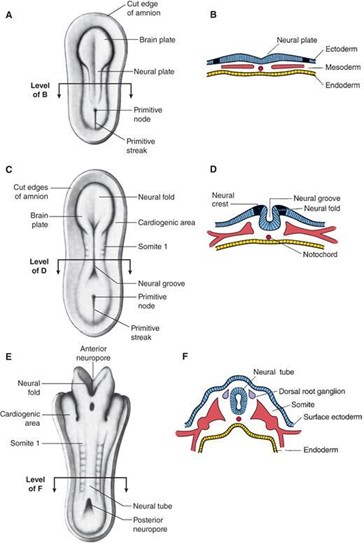
Neurulation refers to the formation and closure of the neural tube. The events of neurulation occur as follows:
The notochord induces the overlying ectoderm to differentiate into neuroectoderm and form the neural plate. The notochord forms the nucleus pulposus of the intervertebral disk in the adult.
The neural plate folds to give rise to the neural tube, which is open at both ends at the anterior and posterior neuropores. The anterior and posterior neuropores connect the lumen of the neural tube with the amniotic cavity.
The anterior neuropore closes during week 4 (day 25) and becomes the lamina terminalis. Failure of the anterior neuropore to close results in upper neural tube defects (NTDs; e.g., anencephaly).
The posterior neuropore closes during week 4 (day 27). Failure of the posterior neuropore to close results in lower NTDs (e.g., spina bifida with myeloschisis).
As the neural plate folds, some cells differentiate into neural crest cells.
The rostral part of the neural tube becomes the adult brain.
The caudal part of the neural tube becomes the adult spinal cord.
The lumen of the neural tube gives rise to the ventricular system of the brain and central canal of the spinal cord.
Neural Crest Cells
The neural crest cells differentiate from cells located along the lateral border of the neural plate, which is mediated by BMP-4 and BMP-7. Neural crest cells undergo a prolific migration throughout the embryo (both the cranial region and trunk region) and ultimately differentiate into a wide array of adult cells and structures
Cranial neural crest cells
The rhombencephalon is divided into eight segments called rhombomeres (R1–R8).
Cranial neural crest cells from R1 and R2 migrate into pharyngeal arch 1 (which also receives neural crest cells from the midbrain area).
Cranial neural crest cells from R4 migrate into pharyngeal arch 2.
Cranial neural crest cells from R6 and R7 migrate into pharyngeal arch 3.
Cranial neural crest cells differentiate into the following adult cells and structures:
Pharyngeal arch skeletal and connective tissue components
Bones of neurocranium
Pia and arachnoid
Odontoblasts (dentin of teeth)
Sensory ganglia of cranial nerve (CN) V, CN VII, CN IX, and CN X
Enteric parasympathetic ganglia of the gut (Meissner and Auerbach; CN X) (from circumpharyngeal neural crest)
Trunk neural crest cells
Trunk neural crest cells extend from somite 6 to the most caudal somites and migrate in a dorsolateral, ventral, and ventrolateral direction throughout the embryo.
Trunk neural crest cells differentiate into the following adult cells and structures:
Melanocytes
Schwann cells
Chromaffin cells of adrenal medulla
Dorsal root ganglia
Sympathetic chain ganglia
Prevertebral sympathetic ganglia
Abdominal/pelvic cavity parasympathetic ganglia.
Clinical considerations. Neurocristopathy is a term used to describe any disease related to maldevelopment of neural crest cells.
Vesicle Development of the Neural Tube
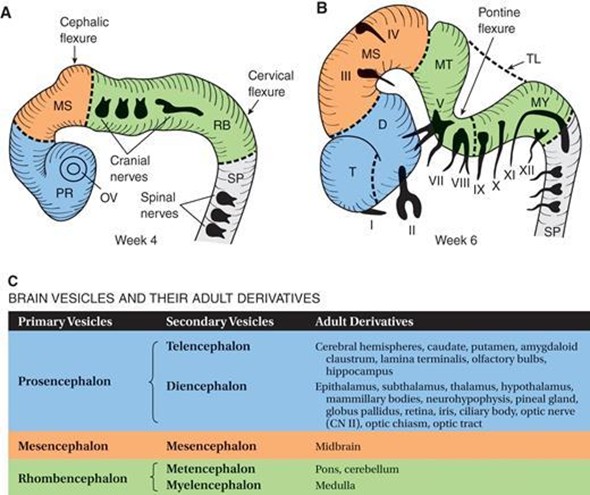
A. Three-vesicle stage of the brain in a 4-week-old embryo. Divisions are indicated by dashed lines. PR 5 prosencephalon (blue); MS 5 mesencephalon (orange); RB 5 rhombencephalon (green); SP 5 spinal cord; OV 5 optic vesicle. B. Five-vesicle stage of the brain in a 6-week-old embryo. Divisions are indicated by dashed lines. Cranial nerves (CN) are indicated by Roman numerals. CN VI is not shown because it exits the brain stem from the ventral surface. T 5 telencephalon; D 5 diencephalon; MS 5 mesencephalon; MT 5 metencephalon; MY 5 myelencephalon; TL 5 tela choroidea; SP 5 spinal cord. C. Table indicating the brain vesicles and their adult derivatives.
The three primary brain vesicles and two associated flexures develop during week 4.
Primary brain vesicles
Prosencephalon (forebrain) is associated with the appearance of the optic vesicles and gives rise to the telencephalon and diencephalon.
Mesencephalon (midbrain) remains as the mesencephalon.
Rhombencephalon (hindbrain) gives rise to the metencephalon and myelencephalon.
Flexures
Cephalic flexure (midbrain flexure) is located between the prosencephalon and the rhombencephalon.
Cervical flexure is located between the rhombencephalon and the future spinal cord.
The five secondary brain vesicles become visible in week 6 of development and form various adult derivatives of the brain.
Telencephalon gives rise to the cerebral hemispheres, caudate, and putamen.
Diencephalon gives rise to the epithalamus, subthalamus, thalamus, hypothalamus, mammillary bodies, neurohypophysis, pineal gland, globus pallidus, retina, iris, ciliary body, optic nerve (CN II), optic chiasm, and optic tract.
Mesencephalon gives rise to the midbrain.
Metencephalon gives rise to the pons and cerebellum.
Myelencephalon gives rise to the medulla.
Histogenesis of the Neural Tube
The cells of the neural tube are neuroectodermal (or neuroepithelial) cells that give rise to the following cell types:
Neuroblasts form all neurons found in the CNS.
Glioblasts are, for the most part, formed after cessation of neuroblast formation. Radial glial cells are an exception and develop before neurogenesis is complete. Glioblasts form the supporting cells of the CNS and include the following:
Astrocytes: project foot processes to capillaries that contribute to the blood–brain barrier, play a role in the metabolism of neurotransmitters (e.g., glutamate, γ-aminobutyrate [GABA], serotonin), buffer the [K1] of the CNS extracellular space, form the external and internal glial-limiting membrane in the CNS, form glial scars in a damaged area of the CNS (i.e., astrogliosis), undergo hypertrophy and hyperplasia in reaction to CNS injury, and contain the glial fibrillary acidic protein (GFAP) and glutamine synthetase, which are good markers for astrocytes.
Oligodendrocytes:produce the myelin in the CNS. A single oligodendrocyte can myelinate several (up to 30) axons.
Ependymocytes(Figure 7.7) line the central canal and ventricles of the brain. These cells are not joined by tight junctions, so that exchange between the cerebrospinal fluid (CSF) and CNS extracellular fluid occurs freely. The drawing (Figure 7.7) shows ependymocytes lining the ventricle of the brain.
Choroid plexus cells(Figure 7.9) are a continuation of the ependymal lining that is reflected over the choroid plexus villi. Choroid plexus cells are modified ependymocytes and secrete CSF by selective transport of molecules from blood coming from the highly vascularized pia mater (called the tela choroidea). These cells are joined by tight junctions (zonula occludens), which are the basis of the blood–CSF barrier.
NOTE: the other glial cells of the CNS are microglia, which are the macrophages of the CNS derived from monocytes and invade the developing nervous system in week 3 along with the developing blood vessels.
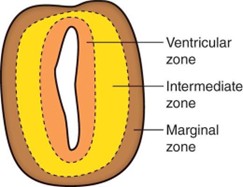
Layers of the Early Neural Tube
Ventricular zone
The early neural tube consists of neuroectoderm arranged in a pseudostratified columnar arrangement.
A first wave of proliferation and differentiation of the neuroectoderm gives rise to neuroblasts, which migrate into the intermediate zone.
A second wave of proliferation and differentiation of the neuroectoderm gives rise to glioblasts, which migrate into the intermediate zone and marginal zone.
The neuroectoderm that remains in the ventricular zone gives rise to ependymocytes, tanycytes, and choroid plexus cells.
Intermediate zone
The intermediate zone contains neuroblasts, which differentiate into neurons with dendrites and axons.
The intermediate zone also contains glioblasts, which differentiate into astrocytes and oligodendrocytes.
The intermediate zone forms the gray matter of the CNS.
The intermediate zone is divided into the alar plate, associated with sensory (afferent) functions, and the basal plate, associated with motor (efferent) functions.
Marginal zone
The marginal zone contains axons from neurons within the intermediate zone.
The marginal zone also contains glioblasts, which differentiate into astrocytes and oligodendrocytes.
The marginal zone forms the white matter of the CNS.
Development of the Spinal Cord
The spinal cord develops from the neural tube caudal to the fourth pair of somites.
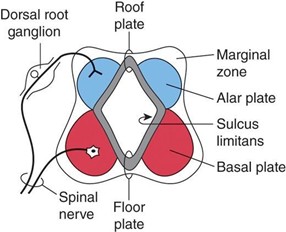
Alar (sensory) plate
The alar plate is a dorsolateral thickening of the intermediate zone of the neural tube.
The alar plate gives rise to sensory neuroblasts of the dorsal horn (general somatic afferent [GSA] and general visceral afferent [GVA] cell regions).
The alar plate receives axons from the dorsal root ganglia that become the dorsal (sensory) roots.
The alar plate becomes the dorsal horn of the spinal cord.
Basal (motor) plate
The basal plate is a ventrolateral thickening of the intermediate zone of the neural tube.
The basal plate gives rise to motor neuroblasts of the ventral and lateral horns (general somatic efferent [GSE] and general visceral efferent [GVE] cell regions).
The basal plate projects axons from motor neuroblasts, which exit the spinal cord and become the ventral (motor) roots.
The basal plate becomes the ventral horn of the spinal cord.
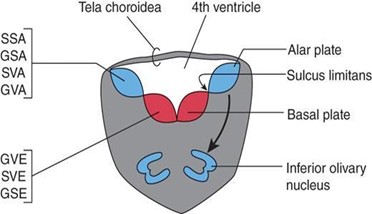
Sulcus limitans (SL)
The SL is a longitudinal groove in the lateral wall of the neural tube that appears during week 4 of development and separates the alar and basal plates.
The SL disappears in the adult spinal cord but is retained in the rhomboid fossa of the brain stem.
The SL extends from the spinal cord to the rostral midbrain.
The roof plate is the nonneural roof of the central canal, which connects the two alar plates.
The floor plate is the nonneural floor of the central canal, which connects the two basal plates. The floor plate contains the ventral white commissure.
Myelination
Myelination of the spinal cord begins during month 4 in the ventral (motor) roots.
Oligodendrocytes accomplish myelination in the CNS.
Schwann cells accomplish myelination in the PNS.
Myelination of the corticospinal tracts is not completed until the end of 2 years of age (i.e., when the corticospinal tracts become myelinated and functional).
Myelination of the association neocortex extends to 30 years of age.
Positional changes of the spinal cord
At week 8 of development, the spinal cord extends the length of the vertebral canal.
At birth, the conus medullaris extends to the level of the third lumbar vertebra (L3).
In adults, the conus medullaris terminates at L1–L2 interspace.
Disparate growth between the vertebral column and the spinal cord results in the formation of the cauda equina, consisting of dorsal and ventral roots (L3–Co), which descends below the level of the conus medullaris.
Disparate growth results in the nonneural filum terminale, which anchors the spinal cord to the coccyx.
Development of the Myelencephalon
SSA = special somatic afferent; GSA = general somatic afferent; SVA = special visceral afferent; GVA = general visceral afferent; GVE = general visceral efferent; SVE = special visceral efferent (aka branchiomeric); GSE = general somatic efferent.
The myelencephalon develops from the caudal rhombencephalon and gives rise to the medulla oblongata.
Alar plate sensory neuroblasts give rise to the following:
Cochlear and vestibular nuclei, which form the special somatic afferent (SSA) column and in the medullopontine junction
Spinal trigeminal nucleus, which forms the GSA column
Solitary nucleus, which forms the special visceral afferent (SVA; taste) and GVA columns
Dorsal column nuclei, which consist of the gracile and cuneate nuclei
Inferior olivary nuclei, which are cerebellar relay nuclei and are derived from the alar plate by migrating to a ventral position
Basal plate motor neuroblasts give rise to the following:
Dorsal motor nucleus of the vagus nerve (CN X) and the inferior salivatory nucleus of the glossopharyngeal nerve (CN IX), which form the GVE column
Nucleus ambiguus, which forms the special visceral efferent (SVE) column (CN IX, CN X, and CN XI)
Hypoglossal nucleus, which forms the GSE column
The roof plate forms the roof of the fourth ventricle. The roof plate is called the tela choroidea, which is a monolayer of ependymal cells covered with pia mater. The tela choroidea is invaginated by pial blood vessels to form the choroid plexus of the fourth ventricle.
The open (rostral) medulla extends from the obex to the stria medullares of the rhomboid fossa. The lateral walls of the rostral medulla open like a book and form the rhomboid fossa (i.e., the floor of the fourth ventricle) due to the formation of the pontine flexure.
Development of the Metencephalon
SSA = special somatic afferent; GSA = general somatic afferent; SVA = special visceral afferent; GVA = general visceral afferent; GVE = general visceral efferent; SVE = special visceral efferent (aka branchiomeric); GSE = general somatic efferent.
The metencephalon develops from the rostral rhombencephalon and gives rise to the pons and cerebellum.
Pons
Alar plate sensory neuroblasts give rise to the following:
Cochlear and vestibular nuclei, which form the SSA column of CN VIII
Spinal and principal trigeminal nuclei, which form the GSA column of CN V
Solitary nucleus, which forms the SVA (taste) and GVA columns of CN VII
Pontine nuclei, which consist of cerebellar relay nuclei (pontine gray) and are derived from the alar plate by migrating to a ventral position (arrow in Figure 7.14)
Basal plate motor neuroblasts give rise to the following:
Superior salivatory nucleus, which forms the GVE column of CN VII
Facial (CN VII) and motor trigeminal (CN V) nuclei, which form the SVE column
Abducent (CN VI) nucleus, which forms the GSE column
Base of the pons. The base of the pons contains the following:
Pontine nuclei from the alar plate
Corticobulbar, corticospinal, and corticopontine fibers, whose cell bodies are located in the cerebral cortex
Pontocerebellar fibers
Cerebellum
The cerebellum is formed from the rhombic lips, which are the two dorsolateral thickened alar plates.
Like the rest of the neural tube, the rhombic lips consist of neuroectoderm arranged in the ventricular zone, intermediate zone, and marginal zone.
In month 3, the neuroectoderm in the ventricular zone undergoes another wave of proliferation to form the internal germinal layer. The internal germinal layer gives rise to the following:
Deep cerebellar nuclei (i.e., dentate, emboliform, globose, and fastigial nuclei)
Purkinje cells
Golgi cells
Some neuroectodermal cells from the internal germinal layer migrate through the marginal zone to form the external germinal layer. The external germinal layer gives rise to the following:
Basket cells
Granule cells
Stellate cells
Both the external and internal germinal layers give rise to astrocytes, Bergmann cells, and oligodendrocytes within the cerebellum
Development of the Mesencephalon
The mesencephalon gives rise to the midbrain.
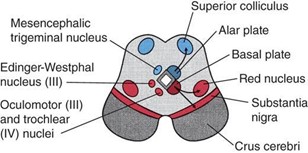
Alar plate sensory neuroblasts gives rise to the superior colliculi and the inferior colliculi (arrows in Figure 7.15 indicate the direction of migration)
Basal plate motor neuroblasts give rise to the following:
Edinger-Westphal nucleus of CN III, which forms the GVE column
Oculomotor (CN III) nucleus, which forms the GSE column
Substantia nigra (arrows indicate direction of migration)
Red nucleus (arrows indicate direction of migration)
Crus cerebri contain corticobulbar, corticospinal, and corticopontine fibers, derived from the cerebral cortex of the telencephalon.
The trochlear (CN IV) nucleus and a portion of the sensory mesencephalic trigeminal (CN V) nucleus originate in the metencephalon and secondarily migrate into the mesencephalon.
Development of the Diencephalon, Optic Structures, and Hypophysis
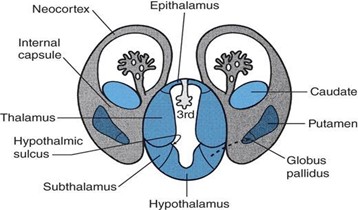
Diencephalon develops from the prosencephalon within the walls of the primitive third ventricle. The alar plates remain prominent in the prosencephalon, but the basal plates regress. The diencephalon gives rise to the epithalamus, thalamus, subthalamus, and hypothalamus.
Epithalamus develops from the alar plate and the embryonic roof plate. The epithalamus gives rise to the following: pineal body (epiphysis), habenular nuclei, habenular commissure, posterior commissure, tela choroidea, and the choroid plexus of the third ventricle.
Thalamus develops from the alar plate. The thalamus gives rise to the following: thalamic nuclei, lateral geniculate body, and the medial geniculate body.
Subthalamus develops from the alar plate. The subthalamus gives rise to the following: subthalamic nucleus, zona incerta, and lenticular and thalamic fasciculi (fields of Fortel). Neuroblasts within the subthalamus migrate (arrow in Figure 7.16) into the telencephalic white matter to become the globus pallidus.
Hypothalamus develops from the alar plate and floor plate ventral to the hypothalamic sulcus. The hypothalamus gives rise to the following: hypothalamic nuclei, mammillary bodies, and neurohypophysis.
Optic vesicles, cups, and stalks are derivatives of diencephalon. They give rise to the retina, iris, ciliary body, optic nerve (CNII), optic chiasm, and optic tract.
Hypophysis (pituitary gland) is attached to the hypothalamus by the pituitary stalk and consists of the anterior lobe (adenohypophysis) and the posterior lobe (neurohypophysis).
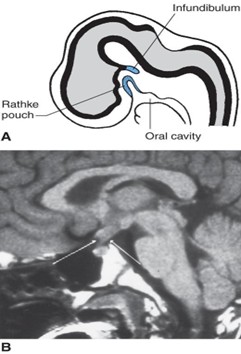
Anterior lobe (adenohypophysis) develops from Rathke’s pouch, which is an ectodermal diverticulum of the primitive oral cavity (stomodeum). The diagram (Figure 17.17A) shows a midsagittal view of an embryo at week 6 indicating Rathke’s pouch and the infundibulum. Remnants of Rathke pouch may give rise to a craniopharyngioma. A craniopharyngioma is the most common supratentorial tumor occurring in childhood and is the most common cause of hypopituitarism in children. The MRI (Figure 7.17B) shows a craniopharyngioma (arrows), which lies suprasellar in the midline, compressing the optic chiasm and hypothalamus.
Posterior lobe (neurohypophysis) develops from the infundibulum, which is a neuroectodermal ventral evagination of the hypothalamus.
Development of the Telencephalon
The telencephalon develops from the prosencephalon. The telencephalon gives rise to the following: cerebral hemispheres, caudate, putamen, amygdaloid, claustrum, lamina terminalis, olfactory bulbs, and hippocampus.
Cerebral hemispheres develop as bilateral evaginations of the lateral walls of the prosencephalic vesicle and contain the cerebral cortex, cerebral white matter, basal ganglia, and lateral ventricles. The cerebral hemispheres are interconnected by three commissures: the corpus callosum, anterior commissure, and hippocampal (fornical) commissure. Continuous hemispheric growth gives rise to frontal, parietal, occipital, and temporal lobes, which overlie the insula and dorsal brain stem. The diagram (at right) shows the development of the cerebral cortex at month 6, month 8, and at term. Note the change in the cerebral cortex from a smooth surface or lissencephalic structure to a convoluted surface or gyrencephalic structure. As growth proceeds, a complex pattern of sulci (grooves) and gyri (elevations) develops.
Cerebral cortex (pallium)
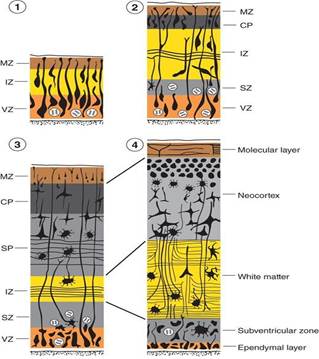
The diagram shows the temporal sequence (early to late: 1–4, respectively) of cytodifferentiation of the cerebral cortex. MZ = marginal zone; IZ = intermediate zone; VZ = ventricular zone; CP = cortical plate; SP = subplate zone; SZ = subventricular zone.
Like the rest of the neural tube, the wall of the telencephalon consists of neuroectoderm arranged in the ventricular zone, intermediate zone, and marginal zone.
Neuroblasts from the ventricular zone and the intermediate zone migrate peripherally to form a transient layer called the cortical plate.
Neuroblasts in the ventricular zone undergo a wave of proliferation to form the subventricular zone.
Neuroblasts from the subventricular zone migrate peripherally to form the subplate zone.
The cortical plate and subplate zone together form the cerebral neocortex.
The intermediate zone becomes devoid of neuroblasts and develops into the white matter of the cerebral hemispheres.
The marginal zone develops into the molecular layer of the cerebral cortex.
The cerebral cortex is classified as the neocortex and allocortex.
Neocortex (isocortex) is a six-layered cortex that represents 90% of the cortical mantle.
Allocortex is a three-layered cortex that represents 10% of the cortical mantle. The allocortex is subdivided into the archicortex (which includes the hippocampal formation) and the paleocortex (which includes the olfactory cortex).
Corpus striatum (striatal eminence)
The corpus striatum appears in week 5 of development in the floor of the telencephalic vesicle.
The corpus striatum gives rise to the basal ganglia: the caudate nucleus, putamen, amygdaloid nucleus, and claustrum.
The corpus striatum is divided into the caudate nucleus and the lentiform nucleus by corticofugal and corticopetal fibers (which make up the internal capsule).
The neurons of the globus pallidus (also a basal ganglion) originate in the subthalamus. These neurons then migrate into the telencephalic white matter and become the medial segments of the lentiform nucleus.
Commissures are fiber bundles that interconnect the two cerebral hemispheres and cross the midline via the embryonic lamina terminalis (commissural plate).
Anterior commissure is the first commissure to appear and interconnects the olfactory structures and the middle and inferior temporal gyri.
Hippocampal (fornical) commissure is the second commissure to appear and interconnects the two hippocampi.
Corpus callosum is the third commissure to appear (between weeks 12 and 22). The corpus callosum is the largest commissure of the brain and interconnects homologous neocortical areas of the two cerebral hemispheres.
Development of the Sympathetic Nervous System
The sympathetic nervous system originates from the basal plate of the neural tube and neural crest cells.
The basal plate of the neural tube gives rise to preganglionic sympathetic neurons within the intermediolateral cell column of the spinal cord, which form white communicating rami found between T1 and L3.
The neural crest cells give rise to postganglionic sympathetic neurons within the sympathetic chain ganglia, prevertebral sympathetic ganglia (e.g., celiac ganglia), and chromaffin cells of adrenal medulla.
Development of the Parasympathetic Nervous System
The parasympathetic nervous system originates from the basal plate of the neural tube and neural crest cells.
The basal plate of the neural tube gives rise to preganglionic parasympathetic neurons within the nuclei of the midbrain (CN III), pons (CNVII), medulla (CN IX, X), and spinal cord at S2–S4.
The neural crest cells give rise to postganglionic parasympathetic neurons within the ciliary ganglion (CN III), pterygopalatine ganglion (CN VII), submandibular ganglion (CN VII), otic ganglion (CN IX), enteric ganglia of the gut (Meissner and Auerbach; CN X), and abdominal/pelvic cavity parasympathetic ganglia.
Development of the Cranial Nerves
Olfactory nerve (CN I) is derived from the nasal (olfactory) placode and mediates smell (olfaction). CN I is capable of regeneration.
Optic nerve (CN II) is derived from the ganglion cells of the retina (which is a diverticulum of the diencephalon) and mediates vision. CN II is not capable of regeneration after transection. CN II is not a true cranial nerve, but a tract of the diencephalon.
Oculomotor nerve (CN III) is derived from the basal plate of the rostral midbrain and mediates eye movements by innervation of the medial rectus muscle, superior rectus muscle, inferior rectus muscle, and inferior oblique muscle, upper eyelid movement by innervation of the levator palpebrae muscle, pupillary constriction by innervation of sphincter pupillae muscle of the iris, and accommodation by innervation of the ciliary muscle.
Trochlear nerve (CN IV) is derived from the basal plate of the caudal midbrain and mediates eye movements by innervation of the superior oblique muscle.
Trigeminal nerve (CN V). The motor division of CN V is derived from the basal plate of the rostral pons. The sensory division of CN V is derived from the cranial neural crest cells. CN V mediates the sensory and motor innervation of pharyngeal arch 1 derivatives.
Abducent nerve (CN VI) is derived from the basal plate of the caudal pons and mediates eye movements by innervation of the lateral rectus muscle.
Facial nerve (CN VII). The motor division of CN VII is derived from the basal plate of the pons. The sensory division of CN VII is derived from the cranial neural crest cells. CN VII mediates the sensory and motor innervation of pharyngeal arch 2 derivatives.
Vestibulocochlear nerve (CN VIII) is derived from the otic placode. The vestibular division of CN VIII mediates balance and equilibrium. The cochlear division of CN VIII mediates hearing.
Glossopharyngeal nerve (CN IX). The motor division of CN IX is derived from the basal plate of the medulla. The sensory division of CN IX is derived from the cranial neural crest cells. CN IX mediates the sensory and motor innervation of pharyngeal arch 3 derivatives.
Vagal nerve (CN X). The motor division of CN X is derived from the basal plate of the medulla. The sensory division of CN X is derived from the cranial neural crest cells. CN X mediates the sensory and motor innervation of pharyngeal arches 4 and 6 derivatives.
Accessory nerve (CN XI) is derived from the basal plate of the spinal segments C1–C6. CN XI innervates the sternocleidomastoid and trapezius muscles.
Hypoglossal nerve (CN XII) is derived from the basal plate of the medulla. CN XIII innervates the intrinsic and extrinsic muscles of the tongue.
Development of the Choroid Plexus
The choroid plexus develops from the roof plates of the rhombencephalon and diencephalon and within the choroid fissure of the telencephalon. The choroid plexus consists of choroid plexus cells (i.e., modified ependymocytes) and a vascular pia mater (tela choroidea). The choroid plexus produces 500 mL of CSF per day. The CSF is returned to the venous system via the arachnoid (granulations) villi of the venous dural sinuses (e.g., superior sagittal sinus).
Congenital Malformations of the Central Nervous System
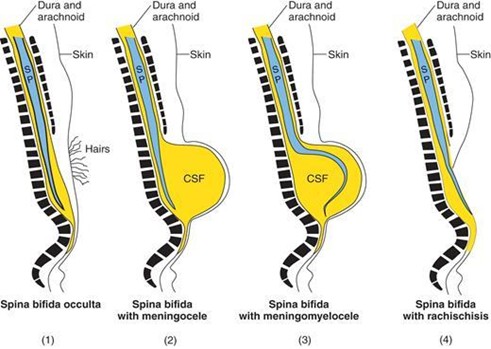
Variations of spina bifida. Spina bifida occurs when the bony vertebral arches fail to form properly, thereby creating a vertebral defect, usually in the lumbosacral region.
SP = spinal cord; CSF = cerebrospinal fluid.
Spina bifida occulta(Figure 7.21) is evidenced by a tuft of hair in the lumbosacral region. It is the least severe variation and occurs in 10% of the population. The MRI (Figure 7.21) of spina bifida occulta shows the presence of the bony vertebral bodies (VB) along the entire length of the vertebral column. However, the bony spinous processes terminate much higher (asterisk) because the vertebral arches fail to form properly. This creates a bony vertebral defect. The spinal cord is intact.
Spina bifida with meningocele(Figure 7.22) occurs when the meninges protrude through a vertebral defect and form a sac filled with CSF. The spinal cord remains in its normal position. The sonogram (Figure 7.22) of spina bifida with meningomyelocele shows the spinal cord (arrows), CSF-filled sac, a small subcutaneous lipoma (L), and the filum terminale (arrowhead).
Spina bifida with meningomyelocele occurs when the meninges and spinal cord protrude through a vertebral defect and form a sac filled with CSF.
Spina bifida with rachischisis(Figure 7.23) occurs when the posterior neuropore of the neural tube fails to close during week 4 of development. This condition is the most severe type of spina bifida, causing paralysis from the level of the defect caudally. This variation presents clinically as an open neural tube that lies on the surface of the back. This condition also falls into a classification called NTDs. Lower NTDs (i.e., spina bifida with rachischisis) result from a failure of the posterior neuropore to close during week 4 of development and usually occur in the lumbosacral region. Upper NTDs (e.g., anencephaly) result from a failure of the anterior neuropore to close during week 4 of development. NTDs can be diagnosed prenatally by detecting elevated levels of α-fetoprotein in the amniotic fluid. About 75% of all NTDs can be prevented if all women capable of becoming pregnant consume folic acid (dose: 0.4 mg of folic acid per day)..
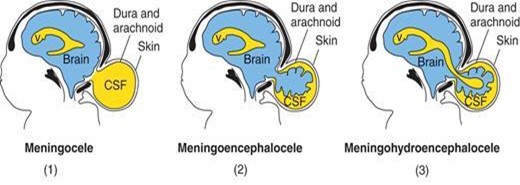
Cranium bifida. Cranium bifida occurs when the bony skull fails to form properly, thereby creating a skull defect, usually in the occipital region.
Schematic drawings illustrating the various types of cranium bifidum.
Cranium bifida with meningocele occurs when the meninges protrude through the skull defect and form a sac filled with CSF.
Cranium bifida with meningoencephalocele occurs when the meninges and brain protrude through the skull defect and form a sac filled with CSF. This defect usually comes to medical attention within the infant's first few days or weeks of life. The outcome is poor (75% of the infants die or have severe intellectual disability).
Cranium bifida with meningohydroencephalocele occurs when the meninges, brain, and a portion of the ventricle protrude through the skull defect.
Anencephaly (meroanencephaly) is a type of upper NTD that occurs when the anterior neuropore fails to close during week 4 of development. This results in the following: failure of the brain to develop (however, a rudimentary brain is present), failure of the lamina terminalis to form, and failure of the bony cranial vault to form. Anencephaly is incompatible with extrauterine life. The photograph (Figure 7.27) shows a newborn infant with anencephaly.
Arnold-Chiari malformation occurs when the caudal vermis and tonsils of the cerebellum and the medulla oblongata herniate through the foramen magnum. This results in a compression of the medulla oblongata and stretching of CN IX, CN X, and CN XII. Clinical features include spastic dysphonia, difficulty in swallowing, laryngeal stridor (vibrating sound heard during respiration as a result of obstructed airways), diminished gag reflex, apnea, and vocal cord paralysis. This malformation is commonly associated with a lumbar meningomyelocele, platybasia (bone malformation of base of skull) along with malformation of the occipitovertebral joint, and obstructive hydrocephalus (due to obliteration of the foramen of Magendie and foramina of Luschka of the fourth ventricle; however, about 50% of cases demonstrate aqueductal stenosis).
The MRI of an Arnold-Chiari malformation. Note the herniation of the brain stem and cerebellum (arrows) through foramen magnum. Note the presence of a fluid-filled syrinx (S) in the cervical spinal cord.
Hydrocephalus is a dilation of the ventricles due to an excess of CSF that may result from either a blockage of CSF circulation or, rarely, an overproduction of CSF (e.g., due to a choroid plexus papilloma). There are two general categories of hydrocephalus:
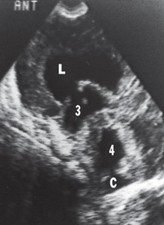
Communicating (or nonobstructive) hydrocephalus. In this type of hydrocephalus, there is free communication between the ventricles and the subarachnoid space. The blockage of CSF in this type of hydrocephalus is usually in the subarachnoid space or arachnoid granulations and results in the enlargement of all the ventricular cavities as well as the subarachnoid space. The sonogram below shows the dilated lateral ventricle (L) communicating through a dilated foramen of Monro with a dilated third ventricle (3) and dilated fourth ventricle (4). The cisterna magna (C) is also shown.
Communicating (or nonobstructive) hydrocephalus.
Noncommunicating (or obstructive) hydrocephalus. In this type of hydrocephalus, there is a lack of communication between the ventricles and the subarachnoid space. The blockage of CSF in this type of hydrocephalus is in the foramen of Monro, the cerebral aqueduct, or the foramen of Magendie/foramina of Luschka and results in the enlargement of only those ventricular cavities proximal to the blockage. There are two types of congenital hydrocephalus, both of which produce a noncommunicating (obstructive) hydrocephalus:
Congenital aqueductal stenosis(Figure 7.30) is the most common cause of congenital hydrocephalus. This type may be transmitted by an X-linked trait, or it may be caused by cytomegalovirus or toxoplasmosis. The sonogram (Figure 7.30) shows dilated lateral ventricles (L), dilated third ventricle (3), but normal-size fourth ventricle (4). Therefore, obstruction at the cerebral aqueduct is presumed.
Congenital aqueductal stenosis.
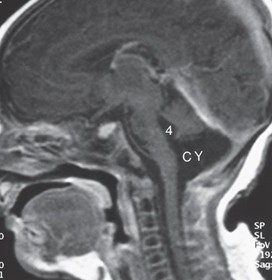
Dandy-Walker syndrome is associated with atresia of the foramen of Magendie and foramina of Luschka (although it remains controversial). This syndrome is usually associated with dilation of the fourth ventricle, posterior fossa cyst, agenesis of the cerebellar vermis, small cerebellar hemispheres, occipital meningocele, and, frequently, agenesis of the splenium of the corpus callosum. The MRI (Figure 7.31) shows a dilated fourth ventricle (4) communicating with a posterior fossa cyst (CY) along with small cerebellar hemispheres.
Dandy-Walker syndrome.
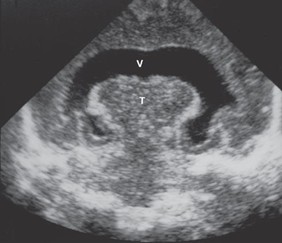
Holoprosencephaly (arhinencephaly; occurs when the prosencephalon fails to cleave down the midline such that the telencephalon contains a single ventricle. It is characterized by the absence of olfactory bulbs and tracts (arhinencephaly) and is often seen in trisomy 13 (Patau syndrome), trisomy 18 (Edward syndrome), short-arm deletion of chromosome 18, and Meckel syndrome. Because the fetal face develops at the same time as the brain, facial anomalies (e.g., cyclopia, cleft lip, cleft palate) are commonly seen with holoprosencephaly. Holoprosencephaly is the most severe manifestation of fetal alcohol syndrome resulting from alcohol use during pregnancy (especially in the first 4 weeks of pregnancy). The sonogram below shows a single, horseshoe-shaped ventricle (V) and fused thalami (T) typical of holoprosencephaly.
Holoprosencephaly (arhinencephaly).
Tethered spinal cord (filum terminale syndrome; Figure 7.36) occurs when a thick, short filum terminale forms. The result is weakness and sensory deficits in the lower extremity and a neurogenic bladder. Tethered spinal cord is frequently associated with lipomatous tumors or meningomyeloceles. Deficits usually improve after transection. The MRI (Figure 7.36) shows a low-positioned spinal cord (arrows) attached to an intraspinal lipoma (L) typical of a tethered spinal cord.
Tethered spinal cord.
Practice Questions
1. Failure of the anterior neuropore to close results in which condition?
2. A neonate presents with a lumbosacral hair tuft but normal neurologic exam. MRI shows absent spinous processes but intact dura and spinal cord. What is the diagnosis?
5. A newborn has poor suckling and appears hypotonic. Imaging shows hypoplasia of the cerebellar hemispheres and vermis. Anomalous development of which of the following MOST LIKELY contributed to this?
6. A 3-month-old infant, previously healthy at birth, is brought to the clinic because of new-onset leg weakness and decreased response to light touch in the lower extremities. MRI of the lumbosacral spine shows a low-lying conus medullaris in continuity with a sacral intraspinal lipoma. What is the diagnosis?